

Coral reefs are seen by humans as tranquil places full of peaceful fish and colourful corals. This, however, is not the way the reef is seen by the many species that inhabit it, the reef is a dangerous, highly cutthroat place where predators are plentiful and competition is fierce. Due to the capabilities and diversity of reef predators and defense mechanisms, many organisms have developed a range of chemicals used both in attack and defense. These chemicals may be as mild as herbivory inhibitors in marine algae or as lethal as neurotoxins produced by Hapalochlaena spp. octopuses or Conus spp. snails. Chemical production may be widespread within a clade such as polyphenol production in marine algae or more species specific such as the venom gland found in the single genus of blennies, Meiacanthus. Marine organisms have developed some very powerful and specialised chemicals that are now used in medicine as well as having other uses for humans. Competition and the requirement to kill prey quickly means that venoms in marine species are far more powerful than similar venoms in terrestrial organisms.
Using chemicals to inhibit predation is a ploy used by both animals and plants/algae in some cases combined with inhibiting physical structure such as spines. In some cases, such as with many algae and sponge species, chemicals used in defense are simply inhibitors and in most cases herbivorous or predatory species have adapted in ways that allow them to feed on chemical producing species. This is a constant evolutionary battle between predator and prey as algae, sponges and other organisms produce stronger inhibitors and herbivores and predators develop methods of overcoming these chemicals. This means that the inhibitory chemicals produced do not stop them from being eaten but rather reduce the number of species able to feed on them. For most sedentary organisms that produce herbivore or predator inhibiting chemicals, these chemicals are secondary metabolites. Secondary metabolites are organic compounds produced for the purpose of greater survivability or fecundity but are not essential to the survival of the organism.
Most plants and algae produce chemical inhibitors that are collectively known as polyphenols. Polyphenols are a group of chemicals that contain more than one phenol group per molecule and are produced by plants and algae as secondary metabolites for the sole purpose of defense. It has been shown that production of polyphenols produced for defensive purposes are not equally distributed throughout the alga and areas that have been damaged, potentially by herbivory, contain elevated concentrations of these chemicals (Pavia and Toth, 2000). This means an area of the alga where predation is occurring contains higher levels of the inhibitory chemicals and is therefore more distasteful for predators. The reason high levels of polyphenols are not produced throughout the individual is because these chemicals require large amounts of energy for production and this reduces the energy put towards growth, repair and reproduction.
There are several toxins produced by marine dinoflagellates produced to inhibit herbivory and, due to bioaccumulation, build up along the food chain and may be harmful and even lethal to humans or other higher order predators. An example of these toxins that is well known to many people, especially fishermen is the ciguatoxin, which causes ciguatera poisoning. Produced primarily by the marine dinoflagellate, Gambierdiscus toxicus, ciguatoxin is 300 times more toxic than the well known tetrodotoxin (TTX) produced by pufferfish (Inoue et al., 2006). Ciguatoxin and scaritotoxin are similar chemicals which open sodium channels in cell membranes affecting mainly neuronal, cardiac and gastrointestinal tissues (Fleming, 2007) resulting in a range of symptoms including nausea, diarrhea, paradoxical temperature reversal (cold objects feel hot and hot objects feel cold), joint and muscle pain and hypotension (Arnold, 2006). Maitotoxin is a toxin produced by the same dinoflagellate species as ciguatoxin and works by keeping calcium channels open in cell membranes causing membrane blebbing and cell death (Estacion and Schilling, 2001). These 3 toxins are found in fish tissue, usually of high order predators such as barracuda, groupers and amberjack and larger individuals contain higher concentrations of the toxins due to the bioaccumulation and inability of these fish to process or excrete the toxin. It is possible that these toxins originally impacted directly on a wider range of herbivores and even low order predators but through evolution, these consumers have built up a resistance to the chemicals.
Bioaccumulation of toxins in shellfish is also common with shellfish used for human consumption all being filter feeders which may feed on dinoflagellates that produce various toxins. Brevetoxin, which causes neurotoxic shellfish poisoning, is chemically very similar to ciguatoxin and behaves in a similar manner, opening sodium channels and therefore having the same symptoms as ciguatera poisoning. Saxitoxin (STX), which causes paralytic shellfish poisoning, has the opposite effect, closing sodium channels causing muscles to relax leading to respiratory failure and possible death in humans as well as other predators (Horizon Foundation Inc., 2003). There is a group of toxins such as pectenotoxins, yessotoxins and dinophysistoxins which inhibit the proteins that control sodium secretion by intestinal cells which causes nausea, vomiting and stomach pains in humans (Horizon Foundation Inc., 2003).
These toxins have varied effects on their direct predators and it has been shown that first order consumers such as copepods may be greatly affected by ingestion of these toxins (Da Costa et al, 2005) as are shellfish and finfish. These effects are magnified with bioaccumulation and many of these toxins are lethal even in small doses to humans.
Sponges contain calcareous or siliceous spicules which provide a minimal amount of defense against predation but like algae, produce a range of secondary metabolites that provide a chemical defense system. Almost 70% of Caribbean sponge species tested by Pawlik et al. (1995) contained secondary metabolites that deterred predatory fish species. In experiments done by Burns et al. (2003), the predatory Klunzinger’s wrasse, Thalassoma klunzingeri, was deterred simply by the presence of endosome and ectosome extracts from various sponge species in food pellets. This suggests the chemicals are not just useful if a predatory takes a bite but they may be able to smell unpalatable chemicals before predation takes place. Species that feed on sponges are often highly specialised, many aquarists are aware of the difficulties in matching spongivorous nudibranchs with their prey items. This is probably due in part to anti-predatory chemical production by sponges that takes a high level of specialization for predatory animals to develop a physiology that allows these chemicals to be metabolized. Sponges produce a wide range of secondary metabolites including alkaloids which are bitter and deter by taste, and turpenoids, a group of chemicals that produce strong aromas also produced by plants such as eucalyptus trees.
Soft corals have a much less potent sting than many other cnidarians and rely more heavily on chemical defenses. Secondary metabolites produced by soft corals are not dissimilar to those produced by sponges, plants or algae with many of the secondary metabolites falling into the same chemical groups as those produced by other organisms. Some soft corals produce several different secondary metabolites; Sinularia flexibilis produces at least three diterpenes specifically for predatory inhibition, flexibilide, sinulariolide and dihydroflexibilide (Aceret et al, 2001). Some of the chemicals produced by these corals not only defend the colony from predation but also have antimicrobial properties and inhibit growth of some infectious bacteria; flexibilide and sinulariolide both exhibit antibacterial properties (Aceret et al, 1998).
Competition for space on the reef is fierce and sedentary organisms such as corals, soft corals, anemones and algae must devise methods for outcompeting rivals. Some species compete by growing tall and shading nearby rivals, some sting competing species with nematocysts while in many species, especially soft corals, this is achieved through production of secondary metabolites which are released into the water column inhibiting the growth of nearby species. This is especially important for larval or juvenile colonies of soft corals as they grow, once they reach larger sizes, shading becomes an important competitive factor. It has been shown that chemicals produced by some algae species can inhibit growth of both algae and invertebrates as well as slow or stop larval development of bivalves (Nelson et al, 2003).
As predators have evolved to allow them to prey on these toxic organisms, some have taken the extra step to make use of these chemicals for their own defense. This is best shown in nudibranchs which feed on toxic sponges and soft corals and move the toxins to their tissue, making them, in turn, toxic and giving them some defense against potential predators. Because toxins are usually specific to particular prey types, this can work to the advantage of other predators, i.e. if a sponge produces a fish specific toxin, nudibranchs are unaffected by the toxin and can use it to defend themselves against fish. In some cases, the toxin is secreted through the skin of the nudibranch and released into the surrounding water giving off an odor which deters predators.
There are several species of echinoderms that possess various forms of toxins and venoms, most are used as defense mechanisms, some in combination with spines. These toxins are unlikely to cause serious harm to humans unless ingested where a number of toxins have been reported to be fatal. Species from the classes Ophiuroidea, Asteroidea, Holothuroidea and Echinoidea have been found to possess venoms including a number of species that can deliver powerful neurotoxins.
One of the best known inhabitants of the reef for all the wrong reasons is the crown-of-thorns starfish, Acanthaster planci, contain venomous spines used for predatory inhibition. The venom found in this species of seastar contains a variety of chemicals including plancinin, an anticoagulant, and plancitoxin, the major lethal ingredient found in the venom (Ota et al, 2006). Effects of the toxin are largely restricted to the cardiovascular system with symptoms including hypotension, swelling and erythema caused by capillary congestion. While the toxin is not lethal or even seriously harmful in humans, the effects are rapid enough that most potential predators would avoid repeated attempts in these seastars. Members of the genus Echinaster are also reported to contain less harmful toxins which are delivered from pits at the base of thorny spines in much the same manner as A. planci. Other members of the class including Plectaster spp. and Solaster spp. are reported to produce more mild toxins which may cause contact dermatitis (Gallagher, 2006).
Well known to most aquarists are the neurotoxins produced by holothuroideans as a defense against predatory fishes which, if released into the water, can cause deaths of numerous fish in an aquarium. The main toxin produced by these animals is holothurin, a chemical stored in various parts of the body and expelled in a few different ways. The main defensive strategies used by sea cucumbers are evisceration and Cuvierian tubule expulsion, where internal organs are expelled often leaving behind a toxic concoction capable of killing all fish within an aquarium. Evisceration can happen in one of two ways, the more common being the expulsion of the respiratory organs, gonads and digestive system which often contain holothurin. The second is the expulsion of the pharynx and part of the lower intestine. In both cases the expelled organs often contain holothurin, enough to kill or at least disable the offending fish predator. Cuvierian tubule expulsion, which is much less common than evisceration, involves the expulsion of the Cuvierian tubules, a mass of sticky, stringy tissue attached to the respiratory organs. While holothurin is not toxic to many invertebrates, the Cuvierian tubules are sticky enough to trap predators not affected by the holothurin. After expulsion of any internal organs, the cucumber has the ability to regenerate itself whilst leaving behind a poisonous decoy for predators. In the aquarium, holothurin is also a potential killer if a cucumber which contains the toxin dies or is damaged. Holothurin can be released into the water by a stressed or dying animal and even in a large aquarium can have a devastating effect on fish and other animals within the aquarium. There are a number of cucumbers that are not potentially dangerous in an aquarium but many popular species such as sea apples, Pseudocolochirus spp., and some popular sand sifters such as Holothuria edulis and H. atra. It is important to research species suitability before adding any of these animals to the aquarium but Holothuria hilla is always a good, safe option when looking for a cucumber to add.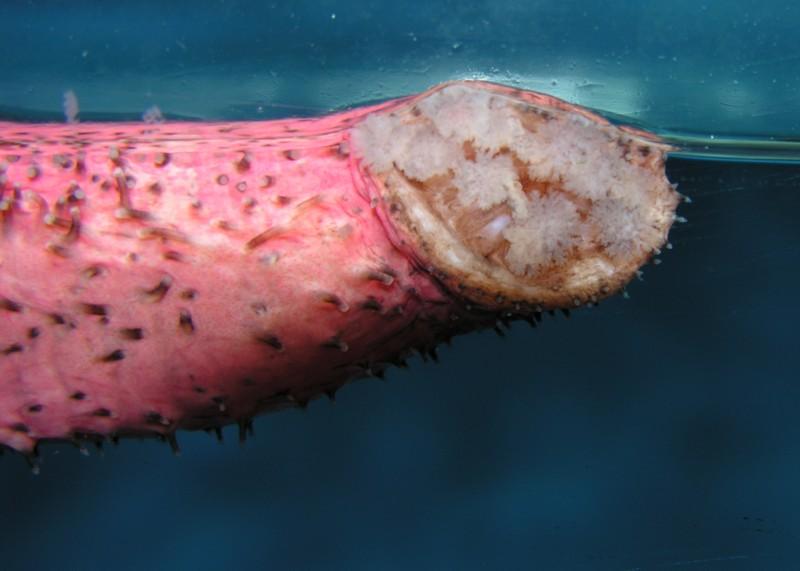
In some species of sea urchins (Echinoidea), various types of chemicals are produced and injected into potential predators via the spines or the pedicellariae which are small beak like structures found on the surface of some echinoderms for defense. In some species such as Tripneustes gratilla, the venom is a protein which breaks up the nearby proteins of the organism that receives the venom (Feigen et al, 1970, cited in Holmes-Farley, 2004). A species of Toxopneustes produces a chemical, Contractin A, which interrupts nervous transmissions and causes muscular contractions. Many species of urchins possess proteinaceous venoms in the tissue covering their spines that inflict minor pain on potential predators. In a more extreme case, the fire urchins such as Asthenosoma varium and A. marisrubri contain venom pouches at the tips of their shorter spines which, when ruptured by pressure, release a venom which causes localized pain and inflammation.
Fish use a myriad of defenses against predation, including toxins and venoms, some produced by the fish themselves, others by symbiotic bacteria. Some fish toxins are harmful, even lethal, to humans if the fish is consumed, others are venomous to touch. An example of this that is well known to aquarists is the venoms found in almost all species of scorpionfishes (Scorpaenidae). While few of these venoms are lethal to humans, most are painful enough to deter potential predators and especially while these fish are young, helps greatly in increasing their chances of survival. While the most common example of these fish are the lionfishes (subfamily Pteroinae), the species that deliver the most harmful dose of neurotoxin is the reef stonefish, Synanceia verrucosa, reportedly responsible for many deaths in tropical island nations (Australian Museum, 2005). In humans, the venom produced by most scorpaenids is not lethal but causes intense pain, edema and necrosis at the site of the sting (Balasubashini et al, 2006). The venom glands are located beneath the dorsal, anal and pelvic fins making it almost impossible for potential predators to consume scorpionfishes whole without receiving a painful sting from one or more of the venomous spines.
Pufferfishes are notorious for their potent toxin, making the fish inedible unless prepared by skilled fugu chefs. The toxin these fish possess is a tetrodotoxin (TTX), named for its presence in fish of the order Tetraodontiformes including pufferfishes and porcupinefishes. This toxin is produced by several bacteria species including Microbacterium arabinogalactanolyticum, Serratia marcescens, Vibrio alginolyticus and Bacillus spp. (Yu et al, 2004; Wu et al, 2005) and is then concentrated in the skin and liver of the fish. TTX is a powerful neurotoxin with symptoms ranging from initial numbness of the tongue and lips to paralysis, seizures and respiratory failure often resulting in death. In humans initial symptoms can occur in as little as 15 minutes and death can be in as little as 4-6 hours (Benzer, 2007). Fish from this family are often brightly coloured, possibly to display their toxic capacity or simply because they are able to display to members of the opposite sex with minimal threat of predation.
Anti-predatory chemicals produced by fish are not always as toxic as they are in scorpionfishes or pufferfishes. A mucous produced by several species of dragonets (Callionymidae) including popular aquarium species, Synchiropus picturatus and S. splendidus, has an unpleasant taste and odor and therefore makes these fish undesirable to predators (Saovy et al, 2005). The dragonet’s mucous, as in all fish species, is derived from a glycoprotein called mucin produced in glandular cells which becomes mucous when mixed with water. There are two cell types which secrete chemicals, the mucous or goblet cells and sacciform cells. The latter are known to produce a range of toxins and distasteful chemicals in various fish species including these few species of dragonets. It is likely the brilliant colours displayed by some of these species acts to warn potential predators of the presence of toxins as this type of colouration is only found in two of the species of dragonets which produce this unpleasant mucous while all species which do not produce these chemicals are more camouflaged. The sacciform cells are also found in several other fish species such as the yellow boxfish, Ostracion cubicus, and believed to produce distasteful chemicals. In the case of the previously mentioned boxfish, the toxin is a haemolytic toxin and while it is distasteful, it also causes breakdown of red blood cells if the fish is consumed.
Non lethal defensive toxins are associated with spines in many fish including rabbitfishes (Siganidae), catfishes (such as Plotosidae) and stingrays. While the combination of injury and venom can be lethal to predators, the toxin itself is not powerful enough to kill most potential predators. In siganids, venom glands are found in grooves within the distal third of the fin rays of the first dorsal, pelvic and anal fins and are capable of delivering a painful sting. In catfishes, the venom is produced in cells surrounding a pair of spines found near the operculum (gill plate) and in some species can inflict a very painful sting. Stingrays produce a neurotoxin from a gland located at the base of the spine which is a modified denticle. In all of these fish, the spine and toxin combination is a powerful deterrent not only to the single attack but also for future attacks on fish of the same species.
An interesting evolutionary adaptation for survival on the reef is shown by the blenny genus Meiacanthus who possess a pair of fangs, enlarged canines, with venom glands at the base. The bite from one of these blennies is no worse than that of a wasp sting but for potential predators, it makes them learn to not attempt to prey on these fish in the future. Bites from these fish are often restricted to the inside of the mouths of fish who have engulfed them but may also be used to ward off attacks from smaller predators unable to take the blenny whole. This successful defense system has lead to the evolution of a variety of mimic species from other genera that use the predators’ knowledge of the venomous Meiacanthus blennies to their advantage.
Venoms are used by many aquatic and terrestrial species to aid in prey capture but those developed by marine species, especially tropical marine species, are often far more toxic than those of comparable terrestrial species. This is due in part to the high level of competition for food; predators must kill and consume their prey quickly in order to ensure their prey is not consumed by a competitor. Venoms are highly diverse and often contain a cocktail or chemicals, some aiding in disabling the prey, others beginning the digestive process.
Blue ring and blue lined octopuses from the genus Hapalochlaena are renowned for their powerful venom which is powerful enough to kill a human by causing paralysis and shutting down the respiratory system. Most people are aware of the toxin called maculotoxin (MTX) that is responsible for a number of human fatalities, but these octopuses also produce a second toxin, hapalotoxin, which is likely to be more useful in prey capture. Maculotoxin is a neurotoxin which causes such symptoms as hypotension and respiratory paralysis (Freeman and Turner, 1970; Trethewie, 1965). It was first thought that MTX and TTX were different toxins with evidence to support the theory they are different chemicals (Croft and Howden, 1972; Savage and Howden, 1977) as well as evidence to suggest they are in fact the same including recent, more conclusive studies (Sheumack et al, 1978; Yotsu-Yamashita et al, 2007). MTX is produced by bacteria, just as TTX is in pufferfish, in the posterior salivary gland of the octopus and is delivered via the beak. This toxin has no effect on crabs and it is therefore likely to be almost exclusively a defense mechanism which causes paralysis and respiratory failure in vertebrates including many of their potential predators such as birds and fish. A secondary use of MTX is the presence of this toxin in the eggs of octopuses from this genus (Sheumack et al, 1984) acting as a predatory inhibitor and increasing the chances of survival of larval octopuses. In humans MTX only causes short term respiratory paralysis and while it can kill quickly, as long as the heart and brain are supplied with oxygen, through CPR or life support, the bite victim will recover with few side effects once the toxin wears off. Victims who survive the first 24 hours usually make a full recovery. The second toxin, hapalotoxin, is more likely to be the toxin used for hunting purposes and is released by the octopus into surrounding water in the vicinity of crabs causing paralysis whereby the octopuses dismembers the crab before consuming it. This allows the octopus to feed on relatively large prey without significant risk of self harm. As with many animal species that possess toxic defenses, Hapalochlaena spp. display bright colours, in this case, flashing bright blue chromatophores in lines or rings to warn potential predators of their venomous nature.
The cone shells of the genus Conus are a highly specialised predator with each of the more than 500 extant species each producing their own form of toxin, known as conotoxins, to target a variety of prey types (Olivera, 1997). Some of the better known and potentially dangerous species such as Conus textilis and C. geographus feed on fish while other species feed on molluscs such as gastropods or bivalves, worms, or even vary their diet as in the case of C. californicus (Kohn, 1966). Because gastropods are by nature slow moving animals, catching faster prey such as small fish could be extremely difficult, this means the toxin must be fast acting and able to immobilize the prey quickly. Anything short of an immediate kill could mean the prey dies a meter from the snail and in a coral reef environment this would simply mean an easy feed for a faster predator. These snails inject their venom via a modified radula which fires harpoon-like projectile which delivers the toxins to the prey. While the ingredients of the venoms are highly varied between species, there are several species of cone shells which produce TTX, many of which feed on small fish and use this powerful neurotoxin to immediately paralyze their prey whereby the fish is consumed whole and still alive.
One of the most dangerous animals to encounter while diving are the almost invisible group predators that include the box jellyfish, Chironex fleckeri and the irukandji jellyfish, Carukua barnesi. This group is the cubozoan jellyfish, highly specialised predators that include species that feed on small fish and are able to disable their prey almost immediately by injecting potent toxins through their nematocysts. While little is known about the chemical makeup of the toxin produced by C. barnesi, it is known that this small (~1”) jellyfish produces a powerful neurotoxin capable of killing humans. The diet of this species includes invertebrates such as small crustaceans in larval animals and larval fishes in mature animals. The powerful toxin enables the fragile jellyfish to catch and paralyze prey quickly and avoid damage to its body. While the toxins of C. fleckeri have also been poorly documented, it is believed the venom produced by this species contains a combination of neurotoxins and myotoxins which cause rapid respiratory and muscular paralysis respectively. Due to the fragile structure of both cubozoan and scyphozoan jellyfish, prey capture must be quick to avoid damage and this is more important for species which prey on larger prey.
Through the dynamics of evolution and the changes in predator-prey relationships, chemicals have come to play an important role in both defense and attack among marine species. With some of the most toxic natural chemicals known to science produced by marine species, predators have become specialised to take advantage of these chemicals, overcome their effects or avoid toxic species. In the aquarium, exposure to these toxins is somewhat uncommon but often very innocuous organisms such as sea cucumbers, urchins or even snails can be potentially dangerous either to the aquarist or the aquarium inhabitants, sometimes both, and care should be taken when selecting new livestock or even when handling fresh live rock.
References
- Aceret, T. L., J. C. Coll, Y, Uchio, P. W. Sammarco. (1998) Antimicrobial activity of the diterpenes flexibilide and sinulariolide derived from Sinularia flexibilis Quoy and Gaimard 1833 (Coelenterata: Alcyonacea, Octocorallia). Comparative Biochemistry and Physiology Part C: Toxicology and Pharmacology. 120 (1):121-126.
- Aceret, T. L., P. W. Sammarco, J. C. Coll, Y. Uchio. (2001) Discrimination between several diterpenoid compounds in feeding by Gambusia affinis. Comparative Biochemistry and Physiology Part C: Toxicology and Pharmacology. 128 (1):55-63.
- Arnold, T. (2006) Toxicity, Ciguatera. http://www.emedicine.com/emerg/TOPIC100.HTM
- Australian Museum (2005). Reef Stonefish. http://www.amonline.net.au/FISHES/students/focus/sverruc.htm
- Balasubashini, M., S. Karthigayan, S. Somasundaram, T. Balasubramanian, P. Viswanathan, V. Menon. (2006) In vivo and in vitro characterization of the biochemical and pathological changes induced by lionfish (Pterios volitans) venom in mice. Toxicology Mechanisms and Methods. 16 (9):525-531.
- Benzer, T. I. (2007) Toxicity, Tetrodotoxin. http://www.emedicine.com/emerg/topic576.htm
- Burns, E., I. Ifrach, S. Carmeli, J. R. Pawlik, M. Ilan. (2003) Comparison of anti-predatory defenses of Red Sea and Caribbean sponges. I. Chemical defense. Mar. Ecol. Prog. Ser. 252:105-114.
- Croft, J. A., M. E. H. Howden. (1972) Toxicon. 10 (6):645-651.
- Da Costa, R. M., J. Franco, E. Cacho, F. Fernández. (2005) Toxin content and toxic effects of the dinoflagellate Gyrodinium corsicum (Paulmier) on the ingestion and survival rates of the copepods Acartia grani and Euterpina acutifrons. Journal of Experimental Marine Biology and Ecology. 322 (2):177-183.
- Estacion, M., W. P. Schilling. (2001) Maitotoxin-induced membrane blebbing and cell death in bovine aortic endothelial cells. BMC Physiology. 1:2.
- Feigen, G. A., L. Hadji, R. A. Pfeffer, G. Markus. (1970) Mode of attack of sea urchin toxin on natural and synthetic substrates. II. Physical properties, substrate specificity, and reaction kinetics of purified fractions. Physiological Chemistry and Physics. 2 (5):427-440.
- Fleming, L. E. (2007) Ciguatera fish poisoning. http://www.whoi.edu/redtide/illness/ciguatera_fish_poisoning.html
- Freeman, S. E., R. J. Turner. (1970) Maculotoxin, a potent toxin secreted by Octopus maculosus Hoyle. Toxicology and Applied Pharmacology. 16 (3):681-690.
- Gallagher, S. A. (2006) Echinoderm Envenomations. http://www.emedicine.com/emerg/topic158.htm
- Holmes-Farley, R. (2004) Sea Urchins: A Chemical Perspective. http://www.reefkeeping.com/issues/2004-08/rhf/index.php
- Horizon Foundation Inc. (2003) Toxic and harmful algal blooms; what you can’t see can hurt you. http://www.bigelow.org/hab/toxin.html
- Inoue, M., K. Miyazaki, Y. Ishihara, A. Tatami, Y. Ohnuma, Y. Kawada, K. Komano, S. Yamashita, N. Lee, M. Hirama. (2006) Total Synthesis of Ciguatoxin and 51-HydroxyCTX3C. J. Am. Chem. Soc. 1280:9352-9354.
- Kohn, A. J. (1966) Food Specialization in Conus in Hawaii and California. Ecology. 47 (6):1041-1043.
- Nelson, T. A., D. J. Lee, B. C. Smith (2003) Are “green tides” harmful algal blooms? Toxic properties of water-soluble extracts from two bloom-forming macroalgae, Ulva fenestrata and Ulvaria obscura. Journal of Phycology. 39 (5):874-879.
- Olivera, B. M. (1996) Conus venom peptides, receptor and ion channel targets and drug design: 50 million years of neuropharmacology. Mol. Biol. Cell. 8 (11):2101-2109.
- Ota, E., Y. Nagashima, K. Shiomi, T. Sakurai, C. Kojima, M. P. Waalkes, S. Himeno. (2006) Caspase-independent apoptosis induced in rat liver cells by plancitoxin I, the major lethal factor from the crown-of-thorns starfish Acanthaster planci venom. Toxicon. 48 (8):1002-1010.
- Pavia, H. and G. B. Toth. (2000). Inducible chemical resistance to herbivory in the brown seaweed Ascophyllum nodosum. Ecology, 81 (11):3212-3225.
- Pawlik, J. R., B. Chanas, R. J. Toonen, W. Fenical. (1995) Defenses of Caribbean sponges against predatory reef fish. I. Chemical deterrency. Mar. Ecol. Prog. Ser. 127:183-194
- Sadovy, Y., J. E. Randall, M. B. Rasotto. (2005) Skin structure in six dragonet species (Gobiesociformes; Callionymidae): interspecific differences in glandular cell types and mucus secretion. Journal of Fish Biology. 66 (5):1411-1418.
- Savage, I. V. E., M. E. H. Howden. (1977) Hapalotoxin, a second lethal toxin from the octopus Hapalochlaena maculosa. Toxicon. 15 (5):463-466.
- Sheumack, D. D., M. E. Howden, I. Spence. (1984) Occurrence of a tetrodotoxin-like compound in the eggs of the venemous blue-ringed octopus (Hapalochlaena maculosa). Toxicon. 22 (5):811-812.
- Sheumack, D. D., M. E. Howden, I. Spence, R. J. Quinn. (1978) Maculotoxin: A neurotoxin from the venom glands of the octopus Hapalochlaena maculosa identified as tetrodotoxin. Science. 199 (4325):188-189
- Trethewie, E. R. (1965) Pharmalogical effects of the venom of the common octopus Hapalochlaena maculosa. Toxicon. 3 (1):55-56.
- Wu, Z., Y. Yang, L. Xie, G. Xia, J. Hu, S. Wang, R. Zhang. (2005) Toxicity and distribution of tetrodotoxin-producing bacteria in puffer fish Fugu rubripes collected from the Bohai Sea of China. Toxicon. 46 (4):471-476.
- Yotsu-Yamashita, M., D. Mebs, W. Flachsenberger. (2007) Distribution of tetrodotoxin in the body of the blue-ringed octopus (Hapalochlaena maculosa). Toxicon. 49 (3):410-412.
- Yu, C., P. H. Yu, P. Chan, Q. Yan, P. Wong. (2004) Two novel species of tetrodotoxin-producing bacteria isolated from toxic marine puffer fishes. Toxicon. 44 (6):641-647.

0 Comments